New approaches to immunotherapy in house dust mite allergy
Article information

Abstract
Allergen immunotherapy (AIT) has developed over the last few decades and has emerged as a promising treatment. House dust mite (HDM) is a target allergen in AIT, and various modified HDM allergens have been improved for their efficacy. Moreover, clinical trials have proved their significantly therapeutic effects in allergy. This article review focuses on HDM allergens developed for AIT efficacy, and determines that their action mechanisms are strongly based on immune tolerance. Treatments for HDM allergens have been optimized by modification or/and addition of adjuvants, and clinically evaluated by subcutaneous and sublingual administration. Representative allergens used in AIT are chemically modified allergoids and recombinant allergens, including altered epitopes. The effective mechanisms subsequent to AIT include the suppressive actions of interleukin-10 and transforming growth factor-β secreted by regulatory cells, including regulatory T cells and regulatory B cells, and isotype switching from immunoglobulin E to immunoglobulin G4. Further studies of AIT are needed to achieve the milestone in treating allergies.
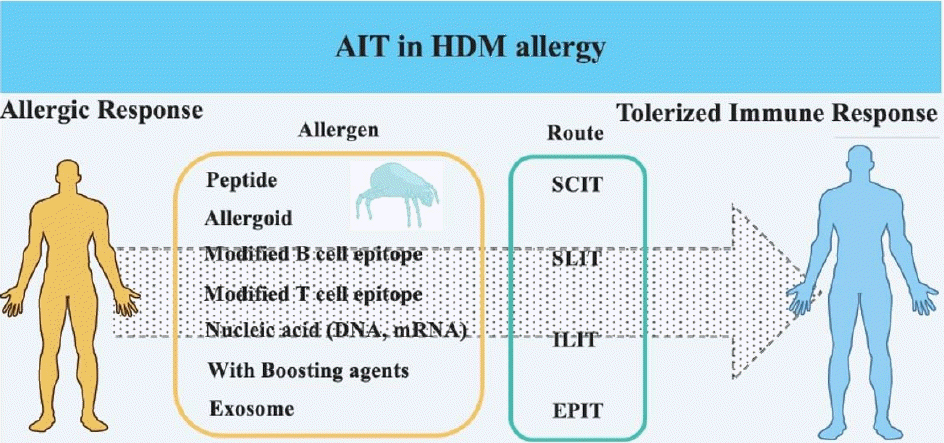
Graphical abstract
Introduction
Before the introduction of allergen immunotherapy (AIT), allergic diseases were considered uncontrolled immune diseases. The first clinical trial of AIT was a game-changer, and AIT has now achieved ground-breaking developments to become an essential treatment for asthma, allergic rhinitis, and atopic dermatitis in children and adults [1]. However, the precise evaluation of its therapeutic mechanism and efficacy has not been accomplished and continues today.
House dust mite (HDM) is a pivotal component of AIT, and new mite allergen proteins are continuously being identified [2,3]. The novel finding of an allergen is accomplished by a draft HDM genome and designed target receptor, as well as classical allergy proteomics [4-7]. Over the last few decades, modified tools for maximizing the use of allergens as powerful vaccines have been developed by unveiling the effective mechanisms related to AIT [8,9]. Allergen structure and form were altered to minimize or decrease allergic responses such as immunoglobulin E (IgE) production and allergic inflammation, practically made by molecular and chemical biology and efficacy of AIT has been finally evaluated. Numerous in vitro and in vivo studies have contributed to the development of AIT for the treatment of allergic disease. In addition, clinical studies performed via different injection routes as well as administration of modified allergen or allergen with adjuvant have revealed that immune tolerance may be achieved by unexpected and complex mechanisms, and there is a need to discover a biomarker for predicting the success of AIT [10].
This review focuses on analyzing the currently available AIT using HDM. Based on the results, we predict the possibility and future of AITs as potential agents for the treatment of allergic diseases.
Allergen types used in AIT
1. Peptides and modified recombinant allergens
Small fragments of allergens are produced by antigen processing and presented by major histocompatibility complex (MHC) in antigen-presenting cells. This step activates T cells and results in allergic inflammation [11-13]. Accordingly, a T-cell-based vaccine has been tested and revealed the possibility of being a useful tool for attenuation of allergy, although the outcome was not remarkable. Since it had no effect on the generation of allergen‐specific immunoglobulin G (IgG) antibodies, clinical trials of this vaccine may be terminated [14,15]. B-cell epitopes are attractive targets for disturbing the allergen-specific IgE production. Although modified B-cell epitopes of Der p 1, Der p 2, and Der p 23 are recognized by B-cell receptors, they cannot induce IgE production but can induce IgG production via different isotype switching [16,17]. In addition, the in silico prediction of T- and B-cell epitopes of Der f 25 may be useful for AIT [18]. Cutting edge of DNA technology and computational structure analysis boosts the antiallergic effects of this vaccine by mutating, aligning, or combining sequences, resulting in the production of mutant, hybrid allergen, or hypoallergen, respectively [19-21].
2. Allergoids
Allergoids are allergens that are chemically modified by formaldehyde and glutaraldehyde. Allergoids lack conformational IgE epitopes but contain linear epitopes of T cells, which are important characteristics for the attenuation of allergic responses. Clinical studies recently demonstrated the usefulness of allergoids in actual patients. This effect is related to the genetic and epigenetic reprograming mechanisms [22,23].
3. Nucleic acid
High doses and repeated injections of an allergen extract elicit unexpected side effects and mortality as well as anaphylaxis. Hence, instead of the protein, DNA and mRNA of an allergen can be recommended as better tools, similar to the coronavirus 2019 vaccine [24,25]. This strategy has been attempted and updated, alongside the development of gene therapy, for patients with gene mutation or deficiency. Such vaccines induce the Th1 type immune response (similar to the immune reaction of nonallergic subjects) and reduce the IgE production induced by Der p 2 or Der p 5 [26,27]. The mRNAs of several allergens were applied to an asthmatic mouse model, and their effects were analyzed and compared. These papers prove that nucleic acid may be beneficial as an effective vaccine material.
4. Conjugation and boosting agents
Adjuvants used for AIT contain aluminum hydroxide, calcium phosphate, and microcrystalline tyrosine. These materials strongly elevate the therapeutic effect of allergoid and modified recombinant allergens [28]. Aluminum hydroxide is a well-known classical adjuvant [29]. Toll-like receptor (TLR) agonists, monophosphoryl lipid A (TLR4 agonist), flagellin A (TLR5 agonist), lipopeptide (TLR2/6 agonist), and cytosine-phosphate-guanosine induce a Th1 response including interferon-γ secretion, which subsequently alleviates the Th2 allergic reaction [30-33]. HDM extract in the liposome protects asthma patients from more severe symptoms and diminishes bronchial provocation due to allergen challenge [34,35]. Liposomes are also used as nucleic acid carriers in DNA or RNA vaccines. The Der p 1 peptide present on virus-like particles efficiently triggers strong IgG responses in human subjects [36]. A recent study reported that virus-like nanoparticles expressing allergens elicited a T helper (Th)1/regulatory T (Treg)-prone response, both in vitro and in an animal model [37]. Liposomes are lipid spheres with the ability to encompass a variety of components, such as therapeutic drugs.
5. Novel approaches
The major AIT using HDM allergen focus on major allergens such as Der p 1, Der p 2, Der f 1, and Der f 2 [38,39]. Der p 5, Der p 7, Der p 10, Der p 21, and Der p 23 are considered as useful components of AIT [40,41]. At the outset, there is a requirement to produce a variety of vaccines for clinical trials using good manufacturing process. This is a pivotal issue that will help to overcome the limited technology available for the development of vaccines required for treating patients afflicted with allergies [42,43]. Moreover, continuous efforts are required to approach newer concepts, in particular, the discovery of a novel tool based on severity of the allergy and personal therapy. Exosomes are small vesicles (10–150 nm) secreted from several types of cells to extracellular environment, and include mRNA, lipids and proteins activating target cell function. Exosomes are arising as promising therapeutic materials in numerous diseases, especially cancer [44,45]. However, exosomes comprise of diverse materials which are as yet unidentified, thereby imparting a disadvantage for its applications. Difficulty in the collection and unknown effective mechanisms of exosomes remain unsolved riddles [46-48]. TLR activators are also used as adjuvants. Der p 38 and Der f 38 has recently been reported as a novel TLR4-binding allergen, which acts as a bidirectional regulator in the transition of predominant eosinophils or/and neutrophils related to asthma severity, and it may be an alternative card in the AIT of HDM allergy [49,50]. A complex combination of current vaccine technologies and an unexpected novel approach will continuously be pursued towards contribution to the development of AIT.
AIT routes
1. Subcutaneous immunotherapy
The most popular route of administering AIT is subcutaneous (SC) injection. Although subcutaneous immunotherapy (SCIT) has been a proven mode for a century, it has several strengths and drawbacks [51]. Its main merit is greater efficacy than other immunotherapies. Its limitations include decreased safety, pain due to repeated injection, longer period for inducing tolerance efficacy, and strong side effects such as severe hypersensitivity. SCIT using HDM has shown maximum efficacy (such as reduction of allergic symptoms and airway hyperreaction and medication use) and preventive effects [52]. Crude HDM extract and modified HDM extract are recommended for treating perennial HDM allergic rhinitis [53]. Recently, an efficacy study of a mouse model evaluated components of the HDM allergen, such as Der p 1 and Der p 2, compared to crude HDM extract [54]. The availability of allergen components will probably help in the future development of SCIT.
2. Sublingual immunotherapy
Sublingual administration is an important and alternative immunotherapy alongside the subcutaneous route. Despite no remarkable efficacy in the treatment for an allergic disease after treatment with HDM extract, sublingual immunotherapy (SLIT) with an HDM tablet reduced allergic symptoms [55-59]. The major merit of SLIT is rare onset of moderate or severe systemic side effects. A meta-analysis has shown that SLIT drops or tablets in HDM immunotherapy are less efficacy than SCIT [60]. Determining the immunotherapy route (SCIT or SLIT) requires numerous factors [61]. To ensure an excellent clinical outcome, clinicians must make a clear decision after considering the efficacy and safety of both procedures as well as patients’ allergy severity. Another important factor is the ability to communicate with the patient, making them psychologically comfortable and ready to continue with regular therapy.
3. Other immunotherapies
Other immunotherapies include oral, intralymphatic immunotherapy (ILIT), and epicutaneous immunotherapy (EPIT). HDM oral immunotherapy in infants with atopy reduces sensitization to allergens; however, no significant preventive effect was noted on HDM sensitization or in symptoms associated with allergy [61].
The ILIT trial was cautiously performed as an alternative method instead of the conventional SCIT and SLIT [62,63]. Allergy symptoms are rapidly alleviated subsequent to ILIT, and its efficacy continues for one year. However, ILIT elicits severe local or systemic hypersensitivity due to the aqueous formulation of allergen extracts used in hypersensitized patients [64]. The safety of cervical ILIT was demonstrated considering that it did not induce moderate to severe adverse side effects [65].
EPIT has been considerably examined in allergic rhinitis. Immunotherapy with microneedles was tested in animal models, and the effective delivery of the HDM extract into the skin was demonstrated [66]. Thus, along with the major methods (SCIT and SLIT), other immunotherapies are steadily being studied for their long-term effects and ability to support better efficacy of immunotherapy.
HDM immunotherapy has been clinically examined to identify more appropriate, efficient, and safe protocols via different injection routes and formulations. HDM proved to be a very useful allergen that imparts protection and prevents allergies (Table 1). There is a steadily growing interest in HDM AIT, and we may need to diversify from familiar methods to create a new concept.
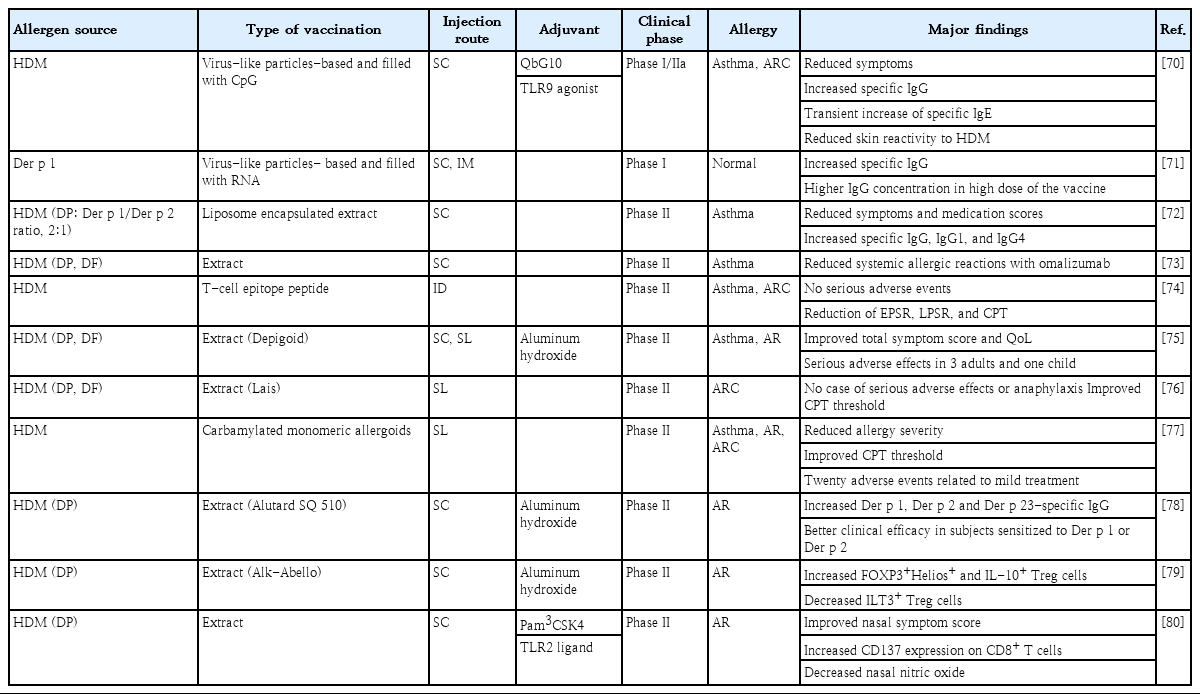
AIT with house dust mite in clinical studies
Effective mechanisms and biomarkers of AIT
1. Immune tolerance
Immune tolerance is a complex and antiallergic mechanism after artificial exposure to a modified allergen [67,68]. First, sensitization to the inhalation or contact of allergen triggers Th2 responses and induces IgE hyperproduction of plasma cells derived from B cells. Basophils and mast cells are finally ready to be activated when a second sensitization occurs. The repetitive and long-term injection of the modified hypoallergen reduces strong allergic responses. The downregulated Th2 cytokines (such as IL-4, IL-5, and IL-13) block the degranulation of basophils and mast cells diminishing the release of histamine and lipid metabolites such as prostaglandin and leukotriene, and decreased tissue injury. Increased levels of IgG4 or IgA prevent the actions of the IgE-mediated allergic cascade accompanied with low IgE levels. The altered isotype switching, i.e., transition of IgE to IgG4, is a pivotal biomarker of AIT outcome (Fig. 1). Novel approaches such as the application of new allergens and exosomes of allergens used in AIT may consolidate the immune tolerance induced by AIT.
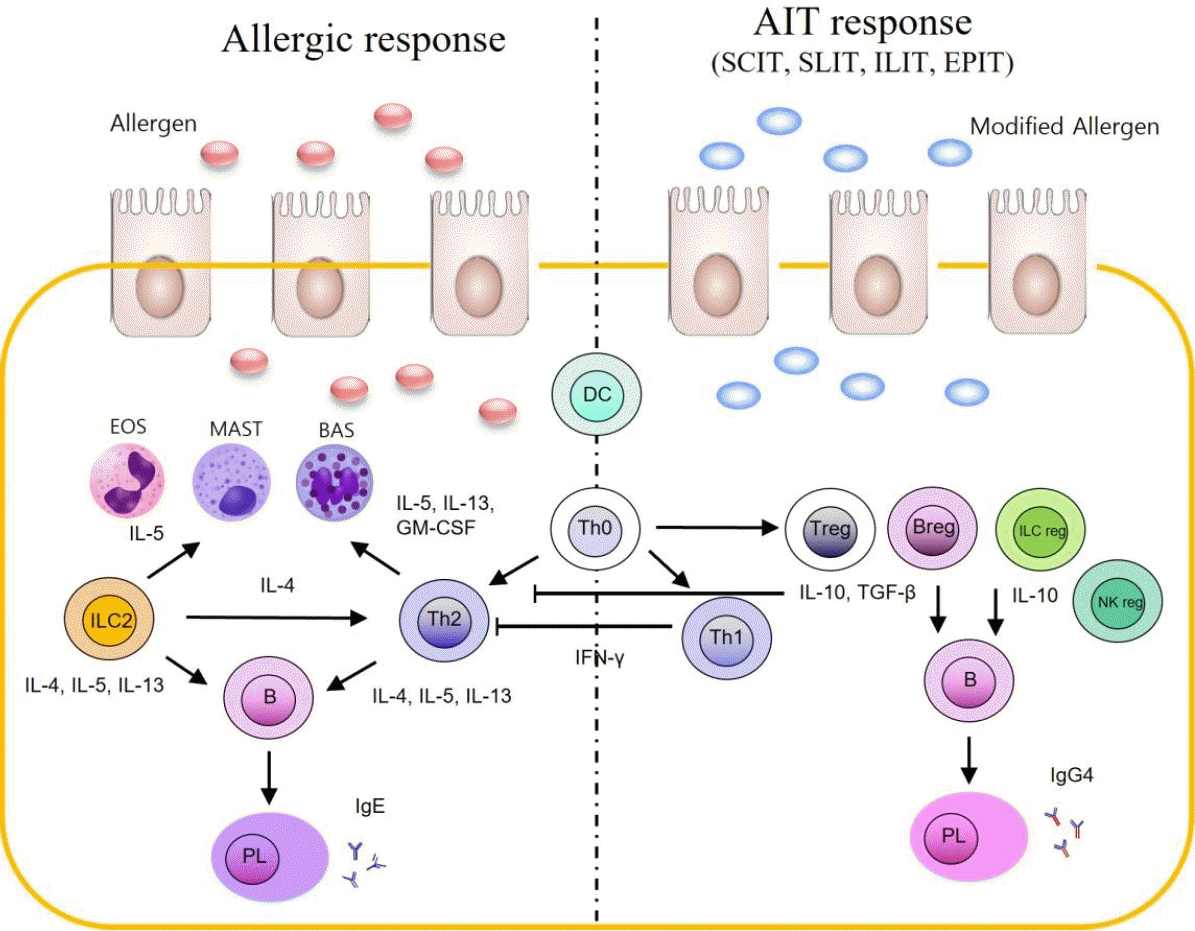
Different allergic mechanisms between naï ve and modified allergens. Onset of allergic response occurs after dendritic cells uptake the allergen, which drives naï ve Th cells (Th0) to be differentiated into Th2 cells. Th2 cytokines such as IL-4, IL-5, and IL-13 induce both eosinophil activation and IgE production of plasma cells differentiated from B cells. Binding of IgE to FcεR activates basophils and mast cells, which subsequently secrete inflammatory mediators. AIT using modified allergen would trigger immune tolerance rather than allergic inflammation. This is caused by modified allergen causing the inhibition of differentiation of Th0 cells to Th2 cells, and activation of regulatory cells. Finally, these complex cascades inhibit allergic inflammation and enhance immune tolerance to the allergen. EOS, eosinophil; MAST, multiple allergen simultaneous test; BAS, basophil; IL, interleukin; GM-CSF, granulocyte-macrophage colony-stimulating factor; ILC, innate lymphoid cell; Th, T helper; IgE, immunoglobulin E; PL, plasma cell; AIT, allergen immunotherapy; SCIT, subcutaneous immunotherapy; SLIT, sublingual immunotherapy; ILIT, intralymphatic immunotherapy; EPIT, epicutaneous immunotherapy; Treg, regulatory T; Breg, regulatory B; NK, natural killer.
2. Dendritic cells
Dendritic cells (DCs) are double-edgeds words, depending on their surroundings. During allergen uptake, DCs function as initiators of allergic inflammation, in which they trigger an immune reaction against bacteria. However, some DCs are able to suppress T-cell activation in a specific situation. DCs consist of 4 essential subtypes: conventional DCs (cDC1s and cDC2s), monocyte-derived DCs, and plasmacytoid DCs. Each subtype is responsible for a variety of immune regulation, including cross-presentation of antigen and Th cell activation. Specifically, tolerogenic DCs are differentiated from immature DCs in the absence of an inflammatory environment, and in the presence of interleukin (IL)-10, transforming growth factor (TGF)-β, or specific stimulators such as lipopolysaccharide, rapamycin, and dexamethasone [69-71]. de Aragão-França et al. demonstrated that tolerogenic DCs lower airway inflammation due to HDM in a mouse model [72]. Sun et al. [73] reported that tolerogenic DCs activate Treg cells in adoptive cell therapy. In case of peripheral tolerance induced by tolerogenic DCs, they are responsible for secreting IL-10 and TGF-β, and these anti-inflammatory cytokines suppress effector T cells and activate Treg cells to accomplish immune tolerance. Moreover, tolerogenic DCs show low expressions of CD80, CD86, and MHC class II unlike other mature DCs. When tolerogenic DCs present the antigen to T cells, the T cells are not activated because no costimulatory signal occurs after the interaction of CD28 with CD80/86 [74,75]. DCs essentially act as linkers for connecting the antigen with bidirectional immune response, and AIT development will be continually required to increase our understanding the regulation of DCs.
3. Regulatory cells
Regulatory cells are derived from T cells, B cells, and natural killer (NK) cells, and are key inducers of immune tolerance or suppressors of allergic responses [51]. IL-10 or/and TGF-β released by regulatory cells inhibits IgE production and increases IgG4 production [76,77]. Conversion from IgE to IgG4 is an inevitable process in immune tolerance after AIT and suppresses the activation of basophils and mast cells due to IgE by kidnapping the allergen.
Treg cells are differentiated from naive T helper cells (Th0 cells) in the absence of costimulatory signal of DCs and presence of TGF-β. CD4+CD25+ Treg1 cells are representative Treg cells that inhibit the Th2 immune response by producing IL-10 and TGF-β on their own [78,79]. CD4+CD25+ forkhead box P3 (FOXP3)+ Treg1 cells act as crucial regulators to achieve efficient HDM immunotherapy. [80] Boonpiyathad et al. [81] suggested the upregulation of Der p 1-specific FOXP3+ Helios+ and IL‐10+ Treg cells, and the downregulation of immunoglobulin‐like transcript 3 (ILT3)+ Treg cells, are associated with improved allergic symptoms after HDM immunotherapy. Programmed cell death1 (PD-1) and cytotoxic T lymphocyte-associated protein 4 are also involved in the immune tolerance mechanism [82,83]. PD-1 modifies the T cells into anergy, as it binds to a counterpart protein PD-ligand 1 of the antigen-presenting cells.
B cells are generally differentiated into plasma cells and secrete IgE antibody against allergens. A different subset of B cells that prefer the inhibition of an excessive allergic response is named regulatory B 1 (Breg1) cells. They are characterized by CD25+ CD71+CD73lowexpression and produce IL-10, TGF-β, and IL-35 to decrease the inflammatory response. IL-10 is a considerably more pivotal anti-inflammatory cytokine than TGF-β and IL-35, based on the evidence that IL-10-overexpressing B cells and highly purified IL-10-producing cells show essential effects of allergen tolerance both in vitro and in vivo [84-86]. IL-10-producing Breg1 cells regulate IgG4 generation and induce Treg cell differentiation, which are the main events required to achieve AIT outcomes [87].
Innate lymphoid cells (ILCs) are classified into ILC1, ILC2, and ILC3, depending on transcription factors and secreted cytokines [88-90]. ILC2 produces IL-4, IL-5, and IL-13 (similar to Th2 cells), and plays a substantial role in the onset and aggravation of allergy due to HDM. Recently, Morita et al. unveiled the existence of IL10-producing regulatory ILCs (ILCregs) originating from ILC2s [90,91].
NK cells are specific lymphocytes in the immune system that act against virus-infected cells and cancer [92]. Distinctive NK cells, revealed by Deniz et al. [93] are responsible for suppressing immune functions by secreting IL-10 and blocking IgE production; these are termed regulatory NK cells. Although NK cells are interesting in the study of allergic diseases, the exact role of regulatory NK cells in AIT remains to be unveiled [92-94].
4. Biomarkers
In line with immune tolerance mechanisms, we can find valuable biomarkers such as increased allergen-specific IgG4, decreased mast cells, eosinophils and ILC2, and increased ILCreg [95]. It has been recently reported that potential gene markers and IL-10 mRNA levels are meaningful biomarkers in HDM AIT study [96,97]. Also, identifying of serum periostin and STAB1 mRNA is invaluable in biomarker development [95]. Biomarkers are highly dependent on AIT tools as well as type of allergies. A single biomarker is insufficient to monitor AIT outcome as a gold standard. According to allergy classification and severity, we must consider the combination of biomarkers and identify novel biomarkers using genomics and proteomics to achieve precise AIT efficacy.
Conclusions
AIT is a promising tool for overcoming allergic diseases such as allergic rhinitis and asthma. However, its AIT application is currently considered challenging and requires further development. There exist several obstacles to fully achieving the purpose of AIT. A novel approach and development of AIT may be required to go overcome 3 categories of drawbacks. First, injection routes and products require to ensure better safety and efficacy. In the case of AIT products, modifications of the allergen and combination of HDM allergen components or/and HDM continue along with thorough quality control. Long-term treatment, painful injections, and side effects are uncomfortable for both patients and clinicians. Second, understanding the mechanism of action would prove invaluable in AIT development. Biomarker development is ongoing for the detection and prognosis of AIT efficacy. Component resolved diagnosis is useful for precision diagnosis and personalized medicine. This challenge needs to be conducted in well-coordinated studies, and will pave the way for AIT success.
Notes
Conflicts of interest
No potential conflict of interest relevant to this article was reported.
Funding
This paper was supported by Eulji University in 2020.