Thyroid peroxidase gene variants and susceptibility to congenital hypothyroidism and autoimmune thyroid disease among Egyptian pediatric cohort
Article information

Abstract
Background
Pediatric thyroid disorders arise from complex interactions among genetic susceptibility, immune dysregulation, and metabolic factors that influence their onset and severity.
Purpose
This study aimed to evaluate the clinical, biochemical, micronutrient, and genetic determinants of thyroid dysfunction in children with particular emphasis on the thyroid peroxidase (TPO) Arg386His and Thr725Pro polymorphisms.
Methods
This case-control study enrolled 150 Egyptian children aged 2–15 years, including 50 with congenital hypothyroidism (CH), 50 with autoimmune thyroiditis (AIT), and 50 controls. The participants underwent clinical assessments, routine laboratory investigations, thyroid function tests, and tests for selenium, copper, and thyroid autoantibodies (Tg Abs and TPO Abs). Following genomic DNA extraction from peripheral blood, the TPO gene polymorphisms (Arg386His and Thr725Pro) were analyzed using polymerase chain reaction-restriction fragment length polymorphism.
Results
Children with thyroid disorders exhibit a higher prevalence of anemia and significantly lower ferritin levels. Profound micronutrient disturbances were evident and characterized by severe selenium deficiency and elevated copper concentrations. The genetic analysis demonstrated a strong association between the TPO exon-8 Arg386His polymorphism and pediatric thyroid disorders. The His allele and His-containing genotypes were significantly more frequent among cases, particularly in children with CH. A genotype-phenotype correlation revealed that the histidine/histidine (His/His) genotype was consistently associated with the highest serumthyroid-stimulating hormone levels in both CH and AIT, suggesting greater functional impairment of thyroid hormone synthesis. Distinct genetic patterns were observed between disease subtypes, with a predominance of the His allele in CH and the Arg allele in AIT. No variation was detected in the Thr725Pro polymorphism, and all participants exhibited the Thr/Thr genotype.
Conclusion
These findings support a multifactorial model of pediatric thyroid disease, with the TPO Arg386His variant, particularly the His/His genotype, emerging as a key genetic contributor to disease susceptibility and severity.
Key message
Question: Which factors influence pediatric thyroid disorder susceptibility and severity?
Finding: Anemia, selenium deficiency, excess copper, and the thyroid peroxidase (TPO) Arg386His polymorphism, especially with the histidine/histidine genotype, are strongly associated with pediatric thyroid disease and high thyroid-stimulating hormone levels.
Meaning: Pediatric thyroid disorders are multifactorial. The TPO Arg386His variant may help identify children at higher risk of severe thyroid dysfunction, enabling earlier diagnosis, improved risk stratification, and more personalized clinical management.
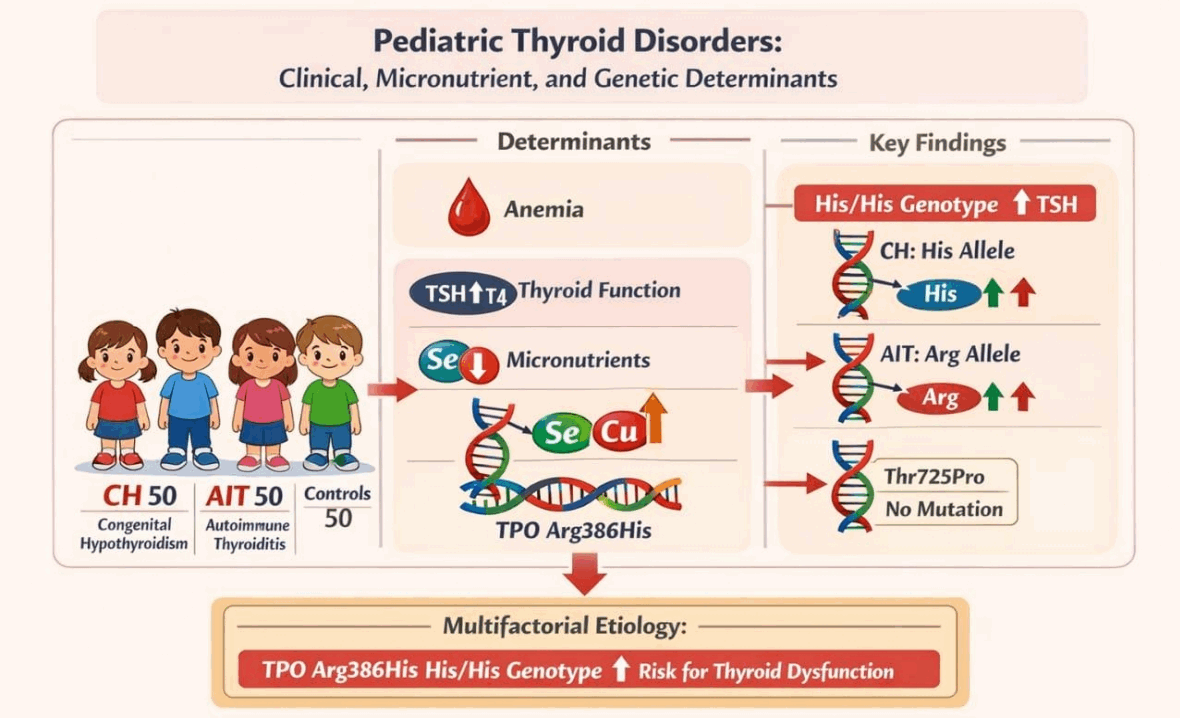
Graphical abstract. CH, congenital hypothyroidism; TSH, thyroid-stimulating hormone; TPO, thyroid peroxidase; His/His, histidine/histidine; AIT, autoimmune thyroiditis; Arg, arginine.
Introduction
Congenital hypothyroidism (CH) arises from developmental or biosynthetic defects of the thyroid gland and represents one of the most preventable causes of intellectual disability when diagnosed and treated early [1]. In contrast, autoimmune thyroiditis (AIT) is the most common cause of acquired hypothyroidism in children and adolescents, characterized by immune-mediated destruction of thyroid follicular cells, accompanied by the production of thyroid autoantibodies, particularly thyroid peroxidase antibodies and antithyroglobulin antibodies [2]. Despite their distinct etiologies—genetic/biosynthetic versus immune-mediated—both disorders ultimately converge on impaired thyroid hormone production, altered metabolic regulation, and adverse effects on growth and development.
Trace elements play an essential role in maintaining normal thyroid function. Iron (Fe) and copper (Cu) are required for optimal thyroid peroxidase (TPO) synthesis and activity, thereby supporting efficient thyroid hormone production [3]. Copper also contributes to redox homeostasis and functions as a cofactor for antioxidant enzymes such as superoxide dismutase, influencing the oxidative environment within thyrocytes [4,5]. Selenium is another critical micronutrient, constituting an integral component of selenoproteins expressed in the thyroid gland, including iodothyronine deiodinases responsible for the conversion of thyroxine (T4) to the biologically active triiodothyronine (T3), as well as glutathione peroxidases that protect thyroid tissue from oxidative damage. Accordingly, selenium deficiency may impair hormone activation and increase susceptibility to autoimmune thyroid dysfunction, and reduced selenium levels have been consistently reported in individuals with autoimmune thyroid disease (AITD) and hypothyroidism [6].
The TPO gene encodes thyroid peroxidase, a thyroid-specific heme-containing enzyme that catalyzes iodide oxidation, iodination of thyroglobulin, and coupling of iodotyrosine residues—key steps in thyroid hormone biosynthesis [7]. Loss-of-function mutations in the TPO gene are a recognized cause of CH due to thyroid dyshormonogenesis, typically inherited in an autosomal recessive pattern and often associated with permanent hypothyroidism and goiter formation [8]. In addition, TPO is a major autoantigen in AITD, and several TPO gene polymorphisms have been investigated for their potential association with susceptibility to autoimmune hypothyroidism. This dual role of TPO—both as a critical enzyme in hormone synthesis and as an immune target—places it at the intersection of genetic and immunemediated thyroid disorders in the pediatric population [9].
Accordingly, the present study aimed to compare children with thyroid disorders (CH and AIT) with healthy controls in terms of demographic, anthropometric, biochemical, micronutrient, and genetic profiles, with particular focus on TPO gene polymorphisms (Arg386His and Thr725Pro).
Methods
1. Study design
A case-control study conducted with 100 Egyptian children and adolescents aged from 2–15 years with thyroid disorders and 50 healthy age and sex-matched control groups. Cases were selected as 50 cases with CH, and 50 cases with AIT. A diagnosis of AIT is made by the demonstration of an elevated concentration of thyroglobulin antibodies and/or thyroid peroxidase antibodies in serum [2]. The study participants were recruited from the pediatric endocrinology clinic and pediatric department of Qena University Hospital, Qena, Egypt, during the study period from May 2024 to October 2025. The study was conducted in accordance with the Declaration of Helsinki. Ethical approval was obtained from the institutional Ethics Committee of the Faculty of Medicine, Qena University (approval code: SVUMEDPED0254251110). Written informed consent was obtained from parents or legal guardians of all participants.
2. Clinical evaluation of the participants
A detailed medical history was obtained for all participants, including age, sex, residence, socioeconomic status, assessed according to Fahmy et al. [10], family history of thyroid disorders, and thyroid-stimulating hormone (TSH) levels at the initial diagnosis. A complete general and systemic clinical examination was performed.
Exclusion criteria included deafness (to exclude Pendred syndrome), Down syndrome, central CH, transient CH, and use of iron or multivitamin supplements within the preceding 6 months.
3. Laboratory and genetic analysis
1) Routine laboratory investigations
Complete blood count, liver enzyme levels, and lipid profile parameters (including triglycerides and total cholesterol) were obtained from the patients’ medical records. Anemia was defined and classified based on age-specific hemoglobin thresholds, as recommended by World Health Organization Guidelines on Hemoglobin Cutoffs to Define Anemia in Individuals and Populations [11]. Serum selenium and copper levels were determined using a commercially available colorimetric assay kit supplied by A.B.C. Diagnostics and Spectrum, Egypt (catalogue Number: 247 001, and 232 001 respectively) and measured by spectrophotometry (Chem-7, Erba, Labomed Inc., Germany). According to the manufacturer’s instructions, age-specific reference ranges were applied for selenium: 0–2 months, 45–90 ng/mL; 3–6 months, 50–120 ng/mL; 7–9 months, 60–120 ng/mL; 10–12 months, 70–130 ng/mL; and >1 year, 70–150 ng/mL. For copper, age- and sex-specific reference ranges were used: infants, 70–140 μg/dL; children (6–12 years), 80–190 μg/dL, adult females, 80–300 μg/dL; and adult males, 70–155 μg/dL. TSH (catalogue Number: MBS2086977, MyBioSource Inc., USA), FT4 (catalogue Number: MBS700784, MyBioSource Inc., USA), FT3 (catalogue Number: MBS262214, MyBioSource Inc., USA), antithyroglobulin (catalogue Number: 89927257, Abnova Thermofisher scientific, USA) and anti-TPO (catalogue Number: MBS4380508, MyBioSource Inc., USA) using commercially available ELISA assays kits. Micro plate ELISA reader EMR-500, USA was used.
2) TPO gene polymorphism analysis [12]
Genetic analysis of the TPO gene focused on 2 singlenucleotide polymorphisms: Arg386His in exon 8 and Thr725Pro in exon 12. Peripheral venous blood samples (2 mL) were collected in EDTA (ethylenediaminetetraacetic acid)-treated tubes, and genomic DNA was isolated using a commercial extraction kit (iNtRON Biotechnology, Korea) in accordance with the manufacturer’s instructions. Extracted DNA was stored at -80°C until further molecular analysis.
Genotyping of both polymorphisms was performed using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) technique. For amplification of the Arg386His variant, the forward primer sequence was 5'-AGA GTC TTA CAA AGG GTG CAC-3' and the reverse primer was 5'-AAG TAC CTG GGA GAG AGA AGC-3'. The Thr725Pro polymorphism was amplified using a forward primer 5'-CTGTCTCGGGTCATCTGTG-3' and a reverse primer 5'-GTA ACG TGG TGT GAG AGG AGAC-3'.
PCR amplification was carried out in a thermal cycler under the following conditions: initial denaturation at 95°C for 5 minutes, followed by 35 cycles of denaturation at 94°C for 45 seconds, annealing at 60°C for 45 seconds, and extension at 72°C for 45 seconds, with a final extension step at 72°C for 5 minutes. The amplified PCR products yielded fragments of 205 bp for the Arg386His polymorphism and 678 bp for the Thr725Pro polymorphism, as confirmed using a 100-bp DNA ladder.
Restriction enzyme digestion of the PCR products was performed at 37°C for 3 hours using FastDigest Cfr42I (SacII) for the Arg386His variant and FastDigest BsaJI (BseDI) for the Thr725Pro variant (Thermo Fisher Scientific, USA). Digested products were separated by electrophoresis on a 2% agarose gel stained with ethidium bromide and visualized under ultraviolet illumination.
For the Arg386His polymorphism, the wild-type arginine/arginine (Arg/Arg) genotype produced 2 fragments of 133 and 76 bp, the heterozygous arginine/histidine (Arg/His) genotype generated 3 fragments of 133, 75, and 58 bp, and the homozygous histidine/histidine (His/His) genotype yielded 2 fragments of 75 and 58 bp (Fig. 1). In the Thr725Pro polymorphism, the Thr/Thr genotype was identified by 3 fragments of 218, 181, and 128 bp (Fig. 2A and B); the Thr/Pro genotype showed 4 fragments of 316, 218, 181, and 128 bp; and the Pro/Pro genotype produced fragments of 316, 181, and 128 bp.

Genotyping of the thyroid peroxidase exon 8 Arg386His polymorphism using polymerase chain reaction-restriction fragment length polymorphism. Restriction enzyme digestion products were resolved by agarose gel electrophoresis. A 100-bp DNA ladder is shown in lane 1. The wild-type Arg/Arg genotype is shown in lanes 2, 3, and 8, characterized by 133- and 76-bp bands. The heterozygous Arg/His genotype is shown in lanes 5, 6, and 7, with 133-, 75-, and 58-bp bands. Homozygous variant His/His genotype is shown in lane 4, displaying 75- and 58-bp bands. Arg/Arg, arginine/arginine; Arg/His, arginine/histidine; His/His, histidine/histidine.
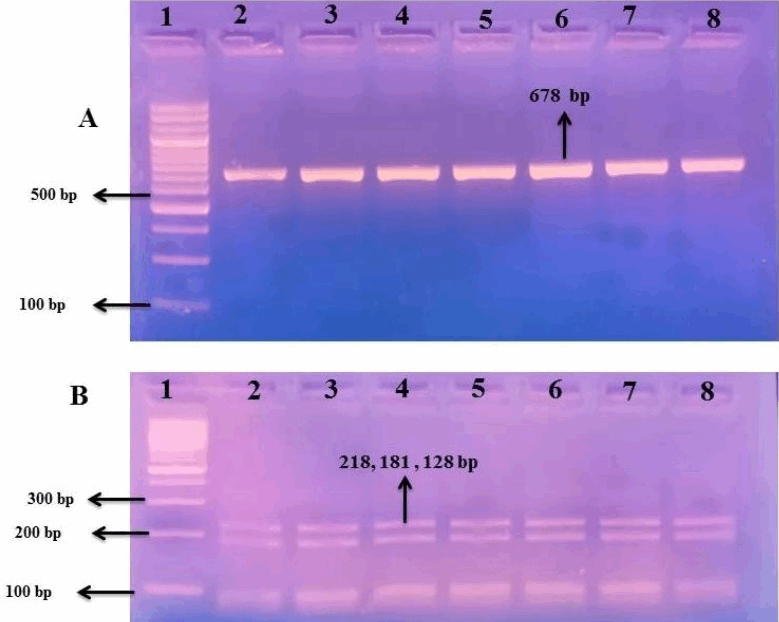
Genotyping of the thyroid peroxidase exon 12 Thr725Pro polymorphism using polymerase chain reaction-restriction fragment length polymorphism. (A) Polymerase chain reaction amplification products showing a single 678-bp band in lanes 2–8, with lane 1 representing a 100-bp DNA ladder. (B) Restriction enzyme digestion products demonstrating the wild-type Thr/Thr genotype in all samples (lanes 2–8), characterized by 218-, 181-, and 128-bp bands. No heterozygous or alternative variant genotypes were detected.
To ensure genotyping accuracy and reproducibility, approximately 10%–15% of the samples were randomly selected and reanalyzed, yielding complete concordance with the initial results.
4. Statistical analysis
Data analysis was conducted using IBM SPSS Statistics ver. 27.0 (IBM Co., USA). Normality testing (Kolmogorov-Smirnov and Shapiro-Wilk tests) indicated non-normal distribution of variables; therefore, data were expressed as median and interquartile range (IQR). Group comparisons were performed using the Mann-Whitney U test or Kruskal-Wallis test with post hoc analysis, as appropriate. Categorical variables were compared using the chisquare test. Multivariate logistic regression analysis with a confidence interval of 95% was used to evaluate the independent risk factors. A 2-tailed P value <0.05 was considered statistically significant. Single nucleotide polymorphisms frequencies were followed the Hardy-Weinberg equation [13].
Results
1. Demographic and clinical data of the studied groups
The demographic and clinical profiles of the studied groups demonstrate overall comparability, with few statistically significant differences. The median age of affected children was 7.4 (IQR, 3.5–10.93) years, similar to controls at 6.5 (IQR, 4–10) years (P=0.806). Sex distribution did not differ significantly, with females comprising 66% of cases and 64% of controls, and males 34% and 36%, respectively (P=0.808). Urban-rural residence was also comparable (urban: 34% of cases vs. 36% of controls; P=0.808), as was socioeconomic status, with most participants in both groups classified as low socioeconomic level (74% vs. 72%, P=0.852). Anthropometric measures showed no significant differences in weight or height between groups. However, body mass index was modestly but significantly higher in children with thyroid disorders (median, 16.7 kg/m²; IQR, 15.46–20.14 kg/m²) compared with controls (median, 15.96 kg/m²; IQR, 14.51–18.23 kg/m²) (P=0.048). Within the case group, 66% had no family history of thyroid disease, while 14% reported a family history of hypothyroidism and 20% of hyperthyroidism.
2. Biochemical and hematological profile of the studied groups
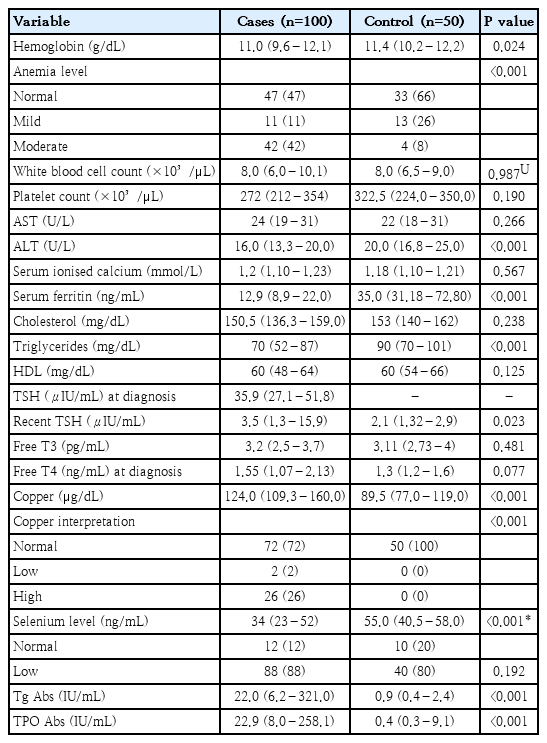
Compared with healthy controls, children with thyroid disorders showed a distinct pattern of hematologic, metabolic, micronutrient, and immunological abnormalities. Hemoglobin levels were significantly lower in cases (median: 11 g/dL vs. 11.4 g/dL), with anemia more prevalent (normal Hb: 47% vs. 66%; moderate anemia: 42% vs. 8%). Iron stores were also reduced, as reflected by lower ferritin concentrations (12.9 ng/mL vs. 35 ng/mL). Total leukocyte and platelet counts were comparable between groups (white blood cell: 8×10³/μL vs. 7.9×10³/μL; platelets: 272×10³/μL vs. 322.5 ×10³/μL). Biochemically, cases had lower alanine aminotransferase levels (16 U/L vs. 20 U/L) a change suggestive of the metabolic suppression characteristic of hypothyroidism rather than hepatic pathology. Lipid profiles were largely similar, although triglycerides were significantly lower in the case group (70 mg/dL vs. 90 mg/dL), reflecting reduced lipid turnover or nutritional insufficiency. Clinically, affected children demonstrated marked thyroid dysfunction at diagnosis, with a median TSH level of 35.88 (IQR, 27.1–51.8) μIU/mL. Recent thyroid function tests demonstrated still higher TSH levels in affected children (3.49 μIU/mL vs. 2.1 μIU/mL in control group), with comparable free T3 (FT3) and free T4 (FT4) concentrations between groups. Marked micronutrient disturbances were observed, with significantly reduced selenium levels in cases (34 ng/mL vs. 55 ng/mL), in contrast, copper levels were significantly elevated (124 μg/dL vs. 89.5 μg/dL). Immunologically, children with thyroid disorders showed pronounced autoimmune activation, with markedly higher TPO antibodies (22.9 IU/mL; IQR, 8.0–258.1 IU/mL) and thyroglobulin antibodies (22.0 IU/mL; IQR, 6.2–321.0 IU/mL) compared with minimal levels in controls, consistent with AIT (Table 1).
Biochemical and hematological profile of children with thyroid disorders versus controls
3. TPO gene polymorphism analysis data in the studied groups
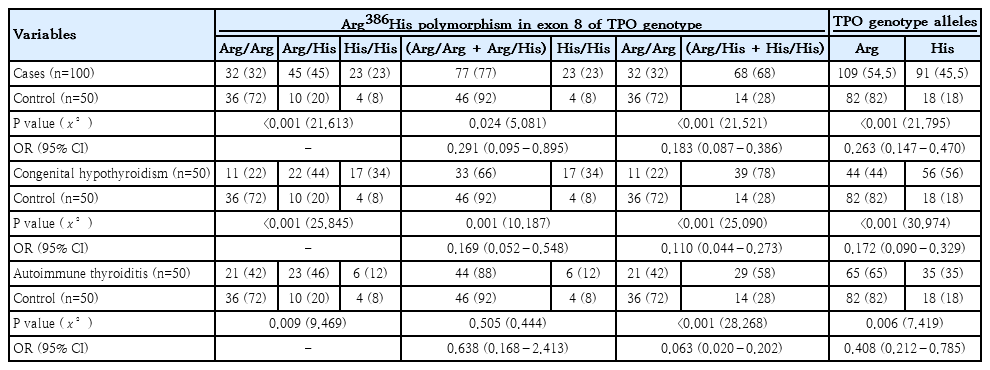
Analysis of the TPO Arg386His (exon 8) polymorphism demonstrated a strong and statistically significant association with pediatric thyroid disorders. In the overall case group, the wild-type Arg/Arg genotype was markedly less frequent than in controls, while the Arg/His and His/His genotypes were substantially overrepresented. This shift toward His-containing genotypes was highly significant (P<0.001) and accompanied by markedly elevated odds ratios, indicating a strong risk effect of the His allele, particularly in homozygous form. This association was even more pronounced in children with CH. The combined variant genotypes (Arg/His+His/His) were present in 78% of CH cases compared with 28% of controls, yielding strong effect estimates with narrow confidence intervals. Correspondingly, the His allele accounted for 56% of alleles in affected children, contrasting sharply with its low frequency among controls.
In AIT, genotype distribution also showed a significant deviation toward variant forms. The Arg/Arg genotype was significantly less common in patients (42%, 21 of 50) than in controls (72%, 36 of 50) (P=0.009, X²=9.469), while the Arg/His genotype was almost twice as frequent in patients (46% vs. 20%). Although the His/His genotype alone did not differ significantly (12% vs. 8%), carriers of the His allele (Arg/His+His/His) were markedly more frequent among patients (58%, 29 of 50) than controls (28%, 14 of 50), with a highly significant association (P<0.001, X²=28.268; odds ratio [OR], 0.063; 95% confidence interval [CI], 0.020–0.202). Allele analysis further confirmed this relationship, with the His allele occurring in 35% of alleles in AIT versus 18% in controls (P=0.006, X²=7.419; OR, 0.408; 95% CI, 0.212–0.785). Collectively, these findings indicate that the Arg386His polymorphism of the TPO gene—particularly carriage of the His allele—is strongly associated with increased susceptibility to both CH and AIT, supporting its role as a genetic risk factor influencing TPO function and thyroid hormone biosynthesis Table 2
Distribution of genotypes and allelic frequencies of TPO Arg386His (exon 8) polymorphism among study cases and subgroups versus controls
The distribution of TPO Arg386His (exon 8) polymorphism shows clear differences between children with CH and those with AIT. In the CH group, the Arg/Arg, Arg/His, and His/His genotypes accounted for 22%, 44%, and 34%, respectively, whereas in AIT these genotypes were distributed as 42%, 46%, and 12%. When genotypes were combined, carriers of at least one Arg allele (Arg/Arg+Arg/His) were significantly less frequent in CH than in AIT (66% vs. 88%). In contrast, the His/His genotype was significantly more prevalent among children with CH (34% vs. 12%). Allelic analysis corroborated these findings, demonstrating a predominance of the His allele in CH (56%), while the Arg allele was more frequent in AIT (65%), with this difference reaching high statistical significance (P=0.003) (Table 3).
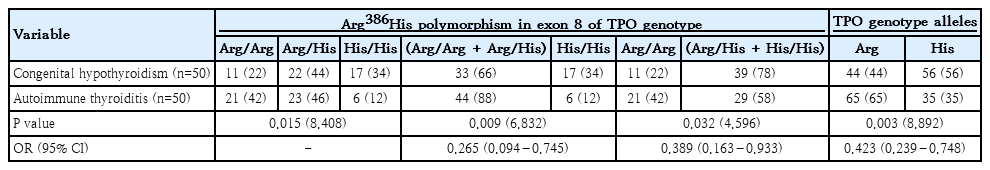
Distribution of genotypes and allelic frequencies of TPO Arg386His (exon 8) polymorphism among congenital hypothyroidism versus autoimmune thyroiditis groups
Serum TSH levels varied significantly according to the TPO exon-8 Arg386His polymorphism across the total cohort of 100 children. Homozygosity for the His allele was associated with the highest TSH concentrations, with a median of 65.2 μIU/mL, compared with substantially lower levels in Arg/Arg (32.24 μIU/mL) and Arg/His (31.57 μIU/mL) genotypes, indicating a strong genotype-phenotype relationship. This association persisted when disease subgroups were analyzed separately. In children with CH, TSH levels were markedly higher in His/His carriers (median, 65.2 μIU/mL) than in those with Arg/Arg (37 μIU/mL) or Arg/His (36.25 μIU/mL), with statistically significant differences in the overall comparison and in pairwise analyses involving the His/His genotype. A similar pattern was observed in AIT, where His/His individuals exhibited the highest TSH values (median, 69.43 μIU/mL), compared with Arg/Arg (25.9 μIU/mL) and Arg/His (27 μIU/mL). Overall, these findings indicate that the His/His genotype of the TPO Arg386His polymorphism is consistently associated with more pronounced TSH elevation at diagnosis in both CH and AIT, supporting a functional impact of this variant on thyroid hormone synthesis efficiency (Table 4).
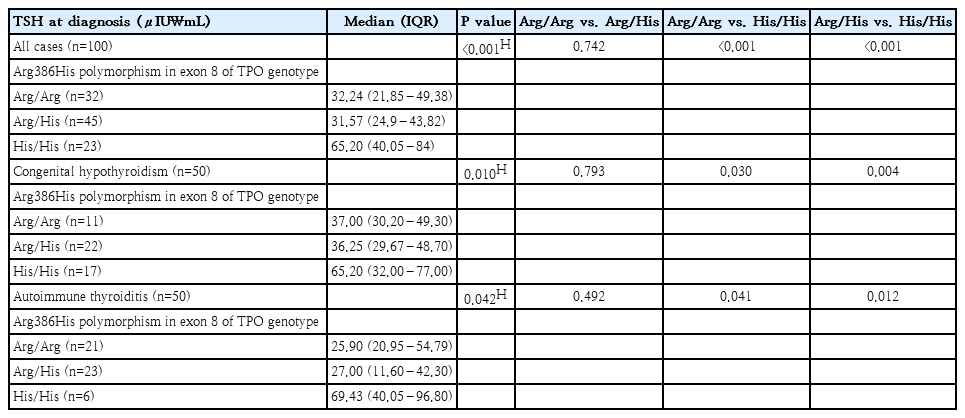
TSH variability across TPO Arg386His genotypes in congenital hypothyroidism versus autoimmune thyroiditis
Genotyping Arg386His polymorphism of exon 8 using RFLP-PCR technique was shown in Fig. 1. As regards Thr725Pro polymorphism of exon 12, all participants were detected to be Thr/Thr genotype as shown in Fig. 2.
In the multivariate binary logistic regression analysis, family history of hypothyroidism emerged as an independent and statistically significant predictor of the studied outcome. After adjustment for other covariates included in the study, individuals with a positive family history of hypothyroidism had a markedly increased likelihood of being His/His genotype carriers, with an adjusted OR of 16.42 (95% CI, 3.35–80.49; P=0.001).
Discussion
The findings from our cohort of children with CH and AIT demonstrate a consistent pattern of hematologic, metabolic, and immunologic disturbances that align closely with the known pathophysiology of thyroid dysfunction and the interplay between nutrition and immune regulation. The significantly lower hemoglobin and ferritin levels observed among affected children indicate that thyroid dysfunction extends beyond an isolated endocrine disorder to substantially influence iron homeostasis and erythropoiesis. Several studies have documented a high prevalence of iron deficiency (ID) in patients with thyroid disorders, particularly hypothyroidism and thyroid autoimmunity, which may impair both the synthesis and biological activity of thyroid hormones [14]. ID adversely affects thyroid hormone production, while thyroid hormone deficiency, in turn, reduces erythroid precursor proliferation both directly and indirectly through diminished renal erythropoietin secretion [15]. Moreover, ID can disrupt the hypothalamic-pituitary-thyroid axis, resulting in altered thyroid hormone levels and reduced responsiveness to TSH. Iron is also a critical cofactor for TPO, the enzyme responsible for catalyzing the iodination of tyrosine residues in thyroglobulin, a key step in thyroid hormone biosynthesis [16].
The trace-element findings in our study highlight a strong association between micronutrient imbalance and thyroid dysfunction. Reduced selenium levels may impair thyroid hormone metabolism and promote autoimmunity, while elevated copper levels likely reflect chronic inflammation and autoimmune activity, as evidenced by increased TPO Abs and Tg Abs. The imbalances in key trace elements—including selenium, copper, iron, zinc, and cobalt—can contribute to thyroid dysfunction through effects on hormone synthesis, oxidative stress, and immune regulation, as emphasized by Zhou et al. [17]. Furthermore, in children and adolescents with AIT, Kyrgios et al. [18] in their randomized trial found that supplementation with organic selenium (200 μg daily) for 6 months significantly reduced antithyroglobulin antibody titers. On the other hand, our demonstration of elevated copper levels in cases (median, ~124 μg/dL vs ~89.5 μg/dL in controls) resonates with evolving evidence on trace-element dysregulation in thyroid disease. Although fewer pediatric data exist, a case-control study conducted by Rostami et al. [19] in adult women with newly-diagnosed Hashimoto’s thyroiditis found that serum copper, selenium, and zinc were significantly different: mean Cu was higher (101.18±33.40 μg/L vs. 82.2±26.82 μg/L, P<0.001) and selenium lower.
Compared with international pediatric cohorts, our findings reveal both shared and distinctive features. Our Egyptian cohort showed more pronounced micronutrient deficiencies and hematologic abnormalities, with concurrent selenium deficiency, ID anemia, and altered copper status, suggesting a compounded effect of nutritional imbalance and inflammation that may exacerbate thyroid dysfunction and autoimmunity.
Thyroid hormones have significant effects on lipid metabolism by regulating hepatic cholesterol synthesis, low-density lipoprotein (LDL)-receptor expression, and lipoprotein clearance. In pediatric patients, both overt and subclinical hypothyroidism have been associated with alterations in lipid profiles, including higher total cholesterol and non–high-density lipoprotein (HDL) cholesterol, which may contribute to early cardiometabolic risk [20,21]. In the current study, no abnormalities were detected in the lipid profiles of the included children, which may be explained by the fact that lipid alterations are influenced by disease severity, duration, and adequacy of levothyroxine therapy; effective treatment generally normalizes lipid parameters. Rivera-Hernández et al. [22] reported that TSH levels are associated with total cholesterol, LDL-C, and HDL-C in hypothyroidism, and that normalization of thyroid function results in reductions in total cholesterol, triglycerides, LDL-C, and overall frequency of dyslipidemia.
Previous studies have identified more than 50 TPO gene mutations resulting from nucleotide deletions, insertions, or sequence substitutions [23]. Such mutations may alter the intracellular localization of the enzyme or disrupt its 3-dimensional structure, leading to impaired enzymatic activity and, consequently, defective thyroid hormone synthesis with subsequent development of thyroid dysfunction [24]. Investigations from multiple countries, including Portugal, Japan, Brazil, and China [25-28], have extensively examined TPO gene mutations in patients with thyroid dyshormonogenesis, with a primary focus on their role in CH. Mutations in the TPO gene are now recognized as a major molecular cause of CH, particularly those affecting exons 8 and 9, which encode the catalytic domain of the enzyme [29]. In agreement with this accumulated evidence, the present study demonstrated an increased frequency of the His allele (Arg/His and His/His genotypes) among affected children, especially in cases of CH, highlighting the pathogenic relevance of TPO gene variability. This observation is further supported by the findings of Arteaga-Jacobo et al. [30], who identified exons 8–9 as mutation hotspots and confirmed TPO defects as a common molecular basis of CH across different populations.
Genetic variation in the TPO gene has also been investigated as a potential susceptibility factor for AITD. Several case-control and population studies have also reported associations between TPO polymorphisms and AITD or altered anti-TPO antibody levels; these reports support the interpretation that nonsynonymous TPO variants can act as susceptibility factors in some populations [31]. Another study conducted by Ahmed et al. [32] found that among Egyptian patients with autoimmune hypothyroidism, SNPs rs2071400 C/T and rs732609 A/C in the TPO gene were significantly more frequent in patients than controls, and correlated with anti-TPO antibody levels. Meanwhile, in a Caucasian (Polish) population, Lacka et al. [33] reported no significant association between certain TPO promoter polymorphisms (rs2071399, rs2071400, rs2071402, rs2071403) and AIT susceptibility, suggesting possible ethnic or regional variation. However, the evidence specifically for Arg386His is not fully investigated. one local study has genotyped Arg386His together with other TPO variants and found different frequencies in patients versus controls; an Egyptian study by Abd El Azeim et al. [12] that included Arg386His in adult cases with autoimmune hypothyroidism, the authors found in patients with AIT, the frequencies of Arg/Arg, Arg/His, and His/His genotypes were 48, 32, and 20%, respectively; and in control participants, the frequencies were 56%, 28%, and 16%, respectively. and suggests population-specific effects may exist.
The Thr725Pro substitution in the TPO protein may alter its secondary structure due to intrinsic chemical differences between threonine and proline. Threonine is a polar, hydrophilic amino acid that can function as a phosphorylation site important for protein activation [34]. In contrast, proline’s rigid cyclic structure imposes conformational constraints on the polypeptide backbone, which can disrupt normal protein folding and destabilize β-helices and β-sheets [35,36]. Additionally, the absence of an amide hydrogen in proline limits its ability to participate in hydrogen bonding, potentially further affecting structural stability [37]. The present pediatric study of CH and AIT revealed no variation in the Thr725Pro polymorphism, as all participants including the control group, exhibited the Thr/Thr genotype. In contrast to our findings, other studies have reported an association between the Thr725Pro polymorphism and thyroid disease in adult populations with autoimmune hypothyroidism. Balmiki et al. [36] and Guria et al. [38] demonstrated that the Pro/Pro genotype was significantly associated with increased disease risk in adults with autoimmune hypothyroidism. This discrepancy highlights that the pathogenic relevance of specific TPO polymorphisms may be age- and population-dependent, with differential effects in pediatric versus adult AITD, underscoring the importance of population-based pediatric genetic studies.
A novel and clinically relevant finding of this study is the differential distribution of TPO Arg386His genotypes between CH and AIT. The His allele predominated in CH, whereas the Arg/His genotype was more frequent in AIT. This divergence suggests that while TPO polymorphisms contribute to susceptibility in both conditions, their pathogenic roles may differ. In CH, the predominance of the His allele—particularly in homozygous form—supports a primary biosynthetic defect in thyroid hormone production. Conversely, in AIT, the higher frequency of Arg-containing genotypes (Arg/His) may indicate that immune-mediated mechanisms, rather than intrinsic enzymatic failure, play a more prominent role, with TPO serving mainly as an autoantigen rather than a functionally deficient enzyme. These findings underscore the genetic heterogeneity of pediatric thyroid disorders and suggest that congenital and autoimmune hypothyroidism represent overlapping but distinct molecular entities. To our knowledge, this is among the first pediatric studies to demonstrate a differential distribution of the Arg386His polymorphism between CH and AIT within the same population, underscoring disease- and population-specific genetic effects.
Despite its strengths, this study has certain limitations. The relatively modest sample size and single-center design may limit generalizability. Future multicenter studies with larger cohorts, functional validation, and long-term follow-up are warranted to further elucidate the mechanistic role of TPO polymorphisms and micronutrient status in pediatric thyroid disease
In conclusion, this study suggests that pediatric thyroid disorders are associated with a complex interaction between genetic susceptibility, micronutrient status, immune activity, and metabolic alterations. The TPO Arg386His polymorphism—particularly the His/His genotype—was significantly associated with increased disease susceptibility and higher TSH levels, indicating a possible relationship with impaired thyroid hormone synthesis. Differences in genotype distribution between CH and AIT further suggest that distinct genetic contributions may exist between these disease entities. The frequent coexistence of selenium deficiency, altered copper levels, and hematological abnormalities highlights the potential value of combined biochemical and genetic assessment in affected children. Collectively, these findings support a more integrated and personalized approach to the evaluation of pediatric thyroid disorders, while underscoring the need for larger, well-powered studies to confirm these associations and explore their clinical implications
Notes
Conflicts of interest
No potential conflict of interest relevant to this article was reported.
Funding
This study received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Acknowledgments
The authors are grateful to everybody who participated in this study; the children who were the participants of this study, the technicians who helped in the laboratory analysis, and the doctors who participated in the collection of the data. Without their help, this study could not have been completed. AI software (ChatGPT) was used only for the graphical abstract, with no role in data analysis or content generation.
Author contribution
Conceptualization: HMS, MHH, SGR, EAA; Data curation: HMS, MHH, AME, MHM, SGR; Formal analysis: HMS, MHH, SGR, AME, MHM; Methodology: MHH, HMS, SGR, EAA; Writing - original draft: HMS; Writing - review & editing: MHH, EAA, SGR